




合作客戶/
拜耳公司 |
同濟大學 |
聯合大學 |
美國保潔 |
美國強生 |
瑞士羅氏 |






相關新聞Info
探索泡沫粗化與表面流變學之間的關聯性疏水性蛋白——結果和討論
來源:上海謂載 瀏覽 1565 次 發布時間:2021-11-25
結果和討論
泡沫穩定性與氣泡尺寸演變
我們已按照上一節所述的程序制備了泡沫,并對所有材料進行了研究。 值得一提的是,HFBII、Quillaja皂甙和b-酪蛋白的發泡溶液含有0.1 wt%的發泡劑,而b-lg的發泡溶液含有0.5 wt%的材料,以達到所需的空氣體積分數。 通過這種方式,我們確保氣泡完全由所選發泡劑以最佳方式穩定。 隨后將這些泡沫轉移到0.5 wt%的黃原膠溶液中,提供足夠高的屈服應力,以防止直徑小于200 mm的氣泡在大約2周內形成泡沫乳狀物(即液體排出)。31最終泡沫樣品含有大約50 vol%的空氣,低于球體的緊密堆積密度, 確保氣泡之間最小的薄膜形成和聚結。 所有這些使我們能夠排除聚結和排水的影響,并將注意力集中在泡沫中氣泡的歧化上。 使用濁度掃描測量設備,我們觀察到所研究系統中隨時間變化的平均氣泡尺寸演變,d(t)/d(0),如圖1所示。 從數據可以清楚地看出,在實驗的時間范圍內(即2周),由HFBII穩定的泡沫沒有顯示出明顯的粗化。 由b-酪蛋白穩定的泡沫粗化速度快得多,約為40分鐘,而b-lg和Quillaja皂甙粗化時間約為70分鐘。 在顯著粗化后,泡沫中的氣泡變得過大,其浮力無法被生物聚合物基質的屈服應力抑制,導致在幾天內形成奶油狀、隨后的聚結和泡沫崩塌。
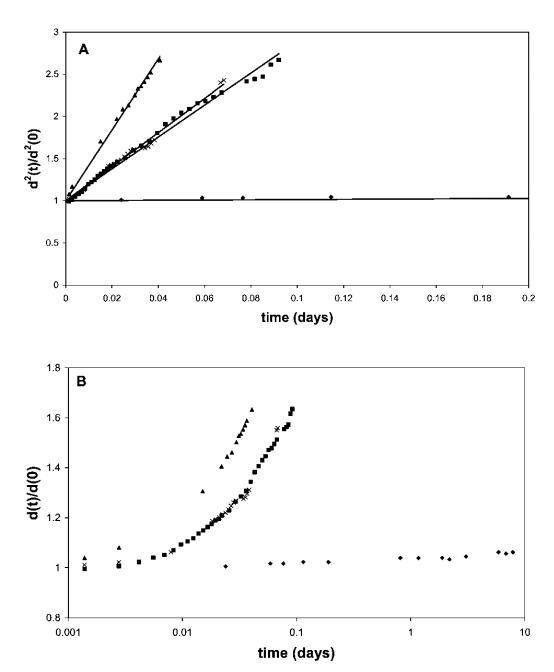
圖1面板A顯示了黃原膠溶液中50 vol%氣泡的相對平方氣泡直徑d2(t)/d2(0)。 泡沫由含有0.1 wt%HFBII(A)、0.5 wt%b-lg(-)、0.1 wt%b-酪蛋白(:)和0.1 wt%Quillaja皂甙(*)的水溶液生成。 將所得泡沫轉移到0.5 wt%黃原膠溶液中。 面板B以對數時間刻度顯示整個時間間隔內的數據。
該結果與先前報道的穩定性試驗結果一致,26表明HFBII穩定的泡沫粗化時間與牛奶蛋白質或皂甙的粗化時間之間存在幾個數量級的差異。 由于泡沫的體積分數低于氣泡的緊密堆積,并且最初抑制了奶油化的影響,因此泡沫穩定性的這些巨大差異歸因于歧化率的差異。 在所有情況下,我們在相同條件下使用相同的氣體成分(空氣)(即氣體溶解度相同),歧化率的差異與空氣/水界面處吸附乳化劑層的表面流變特性有關。 在此背景下,值得一提的是,盡管b-lg和奎拉葉皂甙的泡沫粗化行為非常相似,但奎拉葉皂甙泡沫的整體穩定性(和總壽命)更好。 然而,這應歸因于干燥泡沫中皂甙穩定的薄液膜的更高穩定性,該液膜在后期由于液體排出和大氣泡奶油化而形成。
此外,從這些數據可以清楚地看出,除HFBII外,所有系統的視氣泡半徑都發生了顯著變化,這對應于單個氣泡的空氣/水面處的極大表面變形。 考慮到蛋白質吸附可能是不可逆的,這意味著在歧化發生的大部分時間內,表層處于大變形階段,并且與相的大部分不平衡。 這不同于通常提取表面模量并隨后假設常數的小變形區域。 3,9根據上述觀察和推理,我們繼續研究空氣/水界面擴展層的大變形行為。 考慮到蛋白質吸附可能是不可逆的,這意味著在歧化發生的大部分時間內,表層處于大變形階段,并且與相的大部分不平衡。這不同于通常提取表面模量并隨后假設常數的小變形區域。3,9根據上述觀察和推理,我們繼續研究空氣/水界面擴展層的大變形行為。
大變形膨脹表面流變學
狂犬病HFBII。 圖2a顯示了HFBII的3個壓縮等溫線的典型序列。 曲線中的每個壓縮可分為三個部分。 在低表面壓力下,當蛋白質分子處于非常稀釋的“氣體”狀態時,表面壓力的增加可以忽略不計。 當壓力開始增加,分子失去空間自由度(液相)時,彈性也逐漸增加。 如果層被完全壓縮,則會觀察到曲線中的彎曲點。 考慮到圖中表示表面單層的第一個彎曲點,在約20 mN m-1的表面壓力下,該點處每個HFBII分子的表面積約為400 a?2。 假設球形六邊形緊密堆積,每個分子的面積相當于蛋白質直徑約2.25 nm,這與根據晶體結構確定的分子大小(約2.5 nm)相當好。3
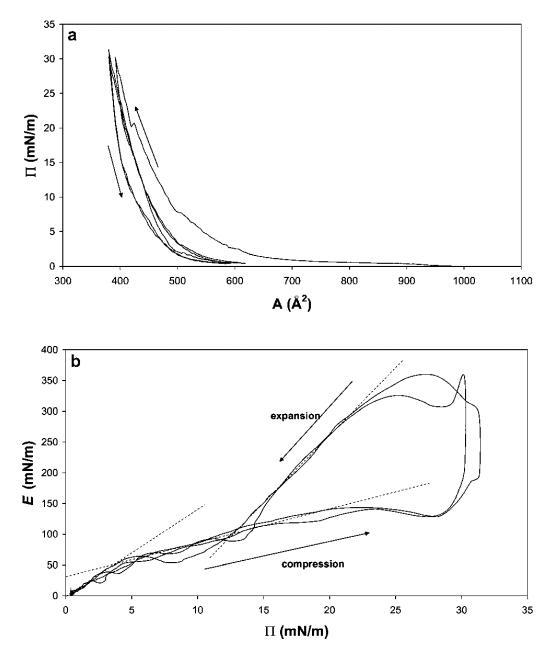
圖2(a)15 mg HFBII在中性pH水上擴散的P–a和(b)E–P曲線,根據相同數據在20℃下測量。 下面板中繪制的線表示不同的狀態。
Paananen等人37測量了HFBII的類似壓力-面積曲線,他們研究了兩種II類疏水蛋白HFBI和HFBII分子膜的結構層次。 報告的工作不同于我們的實驗,因為他們使用pH值為5.0的醋酸鹽緩沖液作為亞相。 與本文給出的結果相比,接近HFBII pI的pH條件可以解釋每個分子的面積略小。
當表面層膨脹時,觀察到明顯的滯后效應,即壓力下降比壓縮時快得多,最后下降到幾mN m-1,與壓縮前類似。 在第二次壓縮時,在與第一次壓縮相似的壓力下,每個分子的面積通常稍小,這表明在第一次壓縮期間分子在表面發生了一些重排。 如果該層被完全壓縮,則可在20 mN m-1的表面壓力周圍看到薄膜屈服。 該屈服點在圖3后面的部分中更為明顯。
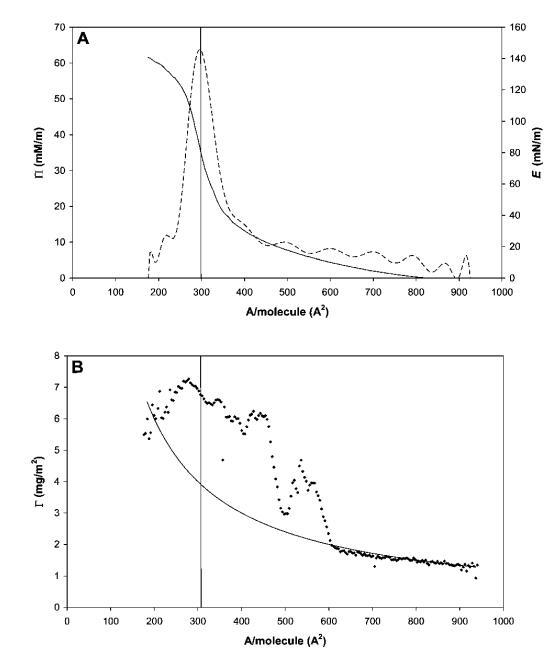
圖3 HFBII的代表性壓縮等溫線,壓縮量遠低于每個分子的面積。 (A) P(-)和E(---)與每個分子的面積的關系。 (B) 攤鋪量和壓縮(Gcomp,-)預期的表面載荷,并通過橢偏儀(Gell,A)測量。
在圖2b中,根據表面壓力P繪制了模量E。可以觀察到三種不同的狀態,與上面討論的類似。 壓縮后,在低P時,觀察到約14的坡度,在Ⅱ=4 mN m-1處,有一個拐點朝向較低的坡度。 該拐點反映了E隨P增加的機制變化。這可能是由HFBII分子形成的松散的自組裝2D網絡引起的,對于弱吸引力粒子也是如此。 第二種狀態持續到20 mN m-1的表面壓力。 在左面板的拐點之外,可以觀察到模量再次開始上升,并具有非常高的斜率。 在這里,2D蛋白質網絡可能已經被壓縮成一個幾乎緊密堆積的單層,其中單個HFBII分子的硬度再次被反映出來。
在屏障方向迅速恢復的點處,觀察到E跳變,這很可能是由于界面適應變形方向變化的時間有限而產生的流體動力效應。 一般來說,經歷短時(高頻)變形的材料沒有時間自然重建/修復其內部結構,這可能導致薄膜破裂。38
隨后,在松弛過程中,模量重新連接到壓縮曲線,表明彈性蒙皮的松弛和重組。 在低于15 mN m-1的壓力下,該網絡可能再次開始可逆地重新排列,以便分解為低于Ⅱ=5 mN m-1的單個實體。
圖3顯示了遠高于單層過渡的代表性壓縮等溫線。 在每分子面積以下進一步壓縮時,表觀表面壓力降至約60 mN m-1,這是非常高的。 該層的強度也反映在143 mN m-1的表觀表面膨脹模量(Eapp)峰值中。 我們必須記住,在這些高表面壓力下,由于表面形成凝膠蛋白質層,表面壓力的測定可能變得不準確。 因此,除了表面流變參數外,圖3B中還提供了橢偏儀數據,這些數據是通過使用實驗部分所述的Multiskop橢偏儀裝置在朗繆爾槽的空氣/水界面進行同步橢偏儀測量獲得的。
橢偏儀數據中,我們可以使用de Feijter的方法在壓縮過程中的每個點獲得蛋白質吸附凝膠(t)量的估計,因為該過程足夠慢,允許我們進行橢偏儀測量。如果蛋白質在整個壓縮過程中仍不可逆地吸附在界面上,則凝膠(t)應根據Gcomp(t)=M0/A(t))遵循表面積的變化,其中M0是界面處蛋白質的初始量,A(t)是槽屏障之間的面積。
如圖3B所示,兩條曲線實際上與表面壓力值一致,約為Ⅱ=5 mN m-1。在這一點之后,Gell突然增加,其值比假設所有蛋白質在界面上均勻分布的估計值高出兩倍。這種行為表明蛋白質層是不均勻的,或者空氣/水界面不是平坦的,這是由于界面上形成了微觀褶皺。事實上,在壓力超過30 mN m-1時,肉眼可以看到界面皺紋的外觀,并且可以在不放大的情況下看到。這些結構的出現開始于移動屏障附近,并隨著壓縮的進行而傳播。為了獲得更好的圖像,我們在表面壓縮和膨脹期間直接進行了光學顯微鏡觀察(圖4),使用底部帶有藍寶石窗口的朗繆爾槽,將其置于透射光學顯微鏡的頂部。如圖5所示的垂直線(在不同放大率下)平行于屏障對齊。
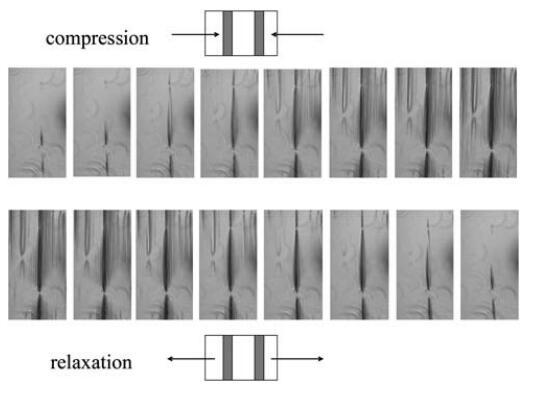
圖4在4倍放大的透射光顯微鏡下,擴展HFBII層的壓縮和松弛。

圖5在透射光顯微鏡下,疏水蛋白在40 mN m-1下的壓縮。左側放大4倍,右側放大10倍。
壓縮時結構的形成和表面積松弛時結構的消失是可逆的,類似于表面性質P、E和凝膠。當在較高放大率下觀察顯微圖像時,觀察到皺紋的均勻空間分布約為4mm。當將橢偏儀數據與顯微圖像進行比較時,我們觀察到橢偏儀信號在比觀察到皺紋的地方低得多的表面壓力下增加。這可能意味著起皺發生在比顯微鏡下觀察到的小得多的長度尺度上。然而,皺紋的初始分布是相當不均勻的,沒有皺紋的區域和皺紋出現的區域具有恒定的空間分布。這種行為與最近發表的Kralchevsky等人的彈性朗繆爾層理論39的預測一致,也可能解釋橢偏測量中的噪聲。
藜蘆皂甙。奎拉葉皂苷的壓縮曲線如圖6所示。該皂甙在拐點處顯示出非常高的表面壓力,最大表面壓力為45 mN m-1。此外,皂甙表現出很寬的遲滯回線。從E–P曲線推斷出斜率為4.3,觀察到E中出現2個平滑的峰,這可能表明界面中存在相變,從皂甙的相對拉長的疏水區域判斷。27當屏障方向逆轉時,觀察到E中存在較大的超調,表明膜破裂,類似于HFBII。
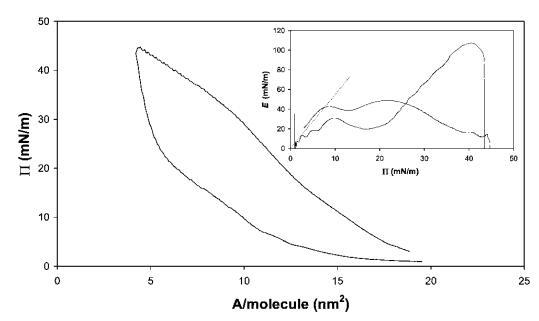
圖6在20℃下測得的30 mg奎拉葉皂苷在中性pH水上的P–A和(插圖)E–P曲線。插圖中繪制的虛線表示E–P曲線的初始坡度。
b-乳球蛋白。圖7顯示了b-乳球蛋白的壓縮等溫線。這條曲線遠沒有HFBII曲線陡峭,拐點更為明顯。盡管在屏障運動逆轉時觀察到滯后現象。E–P曲線的初始斜率為4.3,遠小于低P區的HFBII,表明b-lg分子在界面中相對較軟。觀察到E的平滑最大值為43 mN m-1,之后在進一步壓縮至28 mN m-1時,不再檢測到E的恢復。結果與早期的研究結果一致。40,41在屏障逆轉時,觀察到E的超調,再次表明結構無法適應突然膨脹和斷裂。
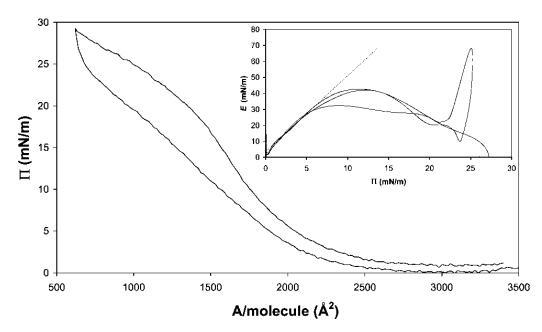
圖7在20℃下測量的15 mg b-乳球蛋白在中性pH水上的P–A和(插圖)E–P曲線。 插圖中繪制的虛線表示E–P曲線的初始坡度。
圖8顯示了被吸附的b-乳球蛋白層的壓縮曲線,在這里,同時進行了橢偏測量。 與HFBII類似,Gell最初遵循Gcomp。 壓縮一段時間后,Gell開始變得比Gcomp小,這表明存在不均勻層或某些材料從表面解吸。 進一步壓縮和膨脹后,凝膠仍小于Gcomp,這表明解吸或表面聚集。 這與其他地方對吸附b-lg層的類似發現一致。42
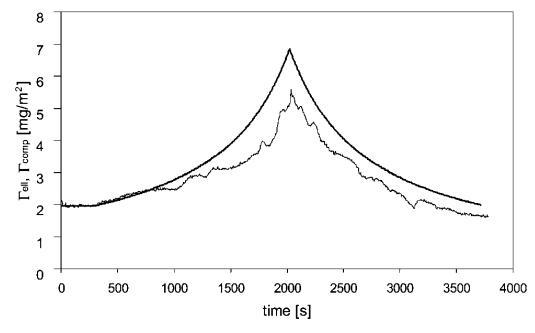
圖8代表性壓縮-吸附b-乳球蛋白(c=0.01 wt%)的膨脹實驗,壓縮低于每個分子的面積。 通過擴散量和壓縮量(Gcomp)和橢偏儀(Gell)測量的預期吸附量。
酪蛋白。 對于純b-酪蛋白,再次觀察到不同的行為(見圖9)。 等溫線的形狀更平滑,沒有觀察到滯后現象。 觀察到的斜率E/P為6.5,與之前報告的吸附層結果一致。43 E–P曲線表明,E的最大值約為25 mN m-1,之后觀察到下降。 該結果與Rodriguez-Patino等人44和Hotrum等人41的早期測量結果一致。此外,E中并未明顯反映出勢壘方向的反轉,這表明結構的松弛速度與勢壘反轉速度一樣快,即約1秒。
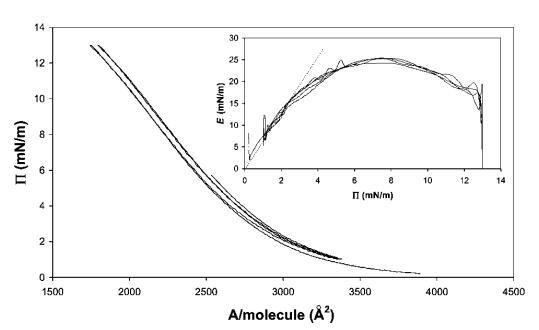
圖9 8 mg b-酪蛋白在水中的P–A和(插圖)E–P曲線,神經pH值,在20℃下測量。 插圖中繪制的虛線表示E–P曲線的初始坡度。
頻率依賴性。 為了評估其中一些系統中粘性損失的重要性,我們對b-lg和HFBII進行了頻率相關的表面膨脹流變。 對于表面膨脹流變學,我們顯示了在18<P<22 mN m-1之間測量的小變形模量(見圖10)。 注意,變形率d(lna)/dt為0.0012 s 1對應于5 mm min 1的勢壘速度,用于大變形實驗。 相比之下,歧化時間約為0.5小時,接近泡沫實驗觀察到的粗化時間。 HFBII的數據表明,在所研究的變形速率范圍內,變形速率對E的影響適中。 對于b-乳球蛋白,頻率的影響更為明顯,因此,在變形速率低于0.001 s 1時,可觀察到E的強烈下降。 這表明b-lg的粘性損失比HFBII更為明顯。 這一比較再次表明,b-lg的外觀更柔軟,壓縮/膨脹時解吸/吸附能力更強。
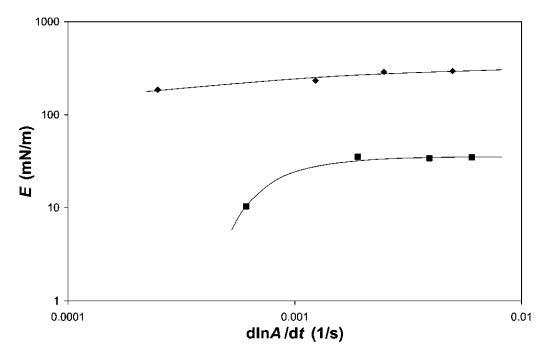
圖10 HFBII(A)和b-lg(-)的表面膨脹模量E,通過18和22 mN m-1之間的線性壓縮-膨脹環測量。 畫線來引導眼睛。
表面剪切流變學。 所有四個系統的表面剪切流變測量如圖11所示。 我們觀察到HFBII具有模量,比b-乳球蛋白和皂甙高一個數量級,這與早期結果一致。25值得一提的是,對于像奎拉葉皂甙這樣的表面活性劑,測得的表面剪切彈性非常高。 b-酪蛋白的表面剪切模量要低一個數量級。 關于頻率,我們發現與HFBII相比,皂甙和b-乳球蛋白對頻率的依賴性更強,尤其是在低頻率下。 此外,對于HFBII而言,Gs 00/Gs0的比率是最低的,即表面彈性對模量的貢獻比表面粘度大得多。 當我們向交叉點外推時,我們預計HFBII和b-乳球蛋白或皂甙之間的弛豫時間會有一個數量級的差異。 對于b-酪蛋白,從0.01 Hz左右的數據可以很容易地觀察到交叉點。

圖11 HFBII(a)、b-乳球蛋白(-)、b-酪蛋白(:)和奎拉葉皂苷(C)的表面剪切儲存(填充符號)和損失模量(開放符號)作為振蕩頻率的函數。
一般性討論
表1列出了從E–P曲線、表面剪切流變模量和泡沫粗化時間獲得的參數匯總。 我們還用冪律擬合了E/P等溫線的初始部分。43這使得我們可以在分子間相互作用可以忽略的(半)稀釋條件下對2D均聚物的行為進行類比,并且使用標度參數可以得出(界面處2D聚合物的)回轉半徑Rg 將線圈中的分段數縮放為Rg f Nn。 結合模量和表面載荷的定義,得出表面壓力和表面膨脹模量E=yP之間的直接關系,其中y與n的關系為n=?y/(y 1)。 對于2D中的自回避線圈,即膨脹聚合物鏈,n等于0.75,而對于Q條件,n等于0.57。 對于折疊線圈,對于折疊分子或硬盤,n進一步降低至0.5。45我們將使用此框架將所考慮的實驗系統的表面行為映射到具有類似膨脹模量與表面壓力依賴關系的等效2D聚合物上。 這將使我們更容易進行比較,并得出一些非瑣碎的結論。
表1在20℃和中性pH條件下進行的膨脹實驗中研究的材料的典型參數。Pmax和Emax表示最大表面壓力和表面膨脹模量。 冪律指數y等于E與P的斜率,n是2D線圈半徑與其分段數的結果縮放指數
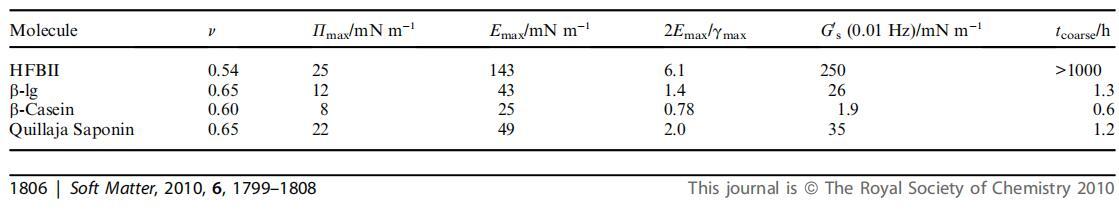
首先,在表觀分子“硬度”方面,可以看出明顯的差異。 HFBII具有更高的斜率y,因此v值明顯較低。 這表明在稀釋狀態下,HFBII表現為等效的緊密折疊聚合物鏈,而牛奶蛋白在中等質量溶劑中表現為等效的2D聚合物鏈。 這與蛋白質結構的差異是一致的。 一方面,HFBII具有相對較小的分子質量和四個二硫鍵,形成一個剛性致密球體,據信在吸附時不會明顯展開。23另一方面,b-lg的分子質量較大,二硫鍵較少。46此外, 經常有人認為,b-lg在吸附時展開。47,48這些特征結合在一起,使分子在空氣-水表面擴散時具有更大的靈活性。
同樣值得一提的是,在這個溫度下,b-lg和b-酪蛋白之間的斜率有一個小但顯著的差異。 雖然這兩個值都表明了q狀態和自回避行走之間的情況,但b-lg聚合物的等效物似乎比b-酪蛋白的自回避程度略高。 在環境溫度下,Hambardzumyan等人認為,b-酪蛋白分子之間的范德華力或氫鍵等吸引性相互作用很重要。 43這可能解釋了與b-lg相比,b-酪蛋白的結構更緊湊,表明b-酪蛋白分子間的相互作用更強, 38因此,由于蛋白質結構的這些差異,不同的蛋白質在壓縮時在表面流變性方面可能表現出很大的差異。 雖然這兩個值都表明了q狀態和自回避行走之間的情況,但b-lg聚合物的等效物似乎比b-酪蛋白的自回避程度略高。在環境溫度下,Hambardzumyan等人認為,b-酪蛋白分子之間的范德華力或氫鍵等吸引性相互作用很重要。43這可能解釋了與b-lg相比,b-酪蛋白的結構更緊湊,表明b-酪蛋白分子間的相互作用更強,這也與其類似于二嵌段共聚物的分子結構相一致。38因此,由于蛋白質結構的這些差異,不同的蛋白質在壓縮時在表面流變性方面可能表現出很大的差異。
最后,等效“聚合物”具有與稀釋奎拉葉皂苷層相同的E/P等溫線,在表面表現為相當軟的實體,這可能是由于其相對較大的每分子面積與分子質量比。27
除了為稀釋表面提供的參數外,還可以從較大壓縮下的數據中獲得更多定性信息。 對于HFBII而言,壓縮后模量持續增加,這可視為一種膨脹硬化。 雖然在單層轉變之后,表觀模量降低,但我們必須意識到,E的下降可能不是真實的。 由于起皺,每個分子的面積不再發生真正的變化,盡管障礙物仍在移動。 此外,由于褶皺,靠近探針的彎月面可能變得不規則,導致P測量中出現偽影。 對于所有其他分子,可以觀察到膨脹軟化,表明軟分子被壓縮或部分分子從表面排出。 因此,各層在壓縮后逐漸變弱。 因此,在歧化過程中的某一點之后,隨著系統阻力的降低,氣泡收縮率將顯著增加。 這與在工藝早期觀察到的情況完全相反,在該過程中,氣泡收縮引起的表面積小幅度減少會導致表面膨脹模量增加,從而加強該層并增加其抗歧化能力。 此外,在大多數理論考慮中,未考慮此類行為,表面模量根據小變形實驗獲得的斜率或使用給定平衡吸附值的吉布斯彈性值作為常數。 對于HFBII而言,壓縮后模量持續增加,這可視為一種膨脹硬化。雖然在單層轉變之后,表觀模量降低,但我們必須意識到,E的下降可能不是真實的。由于起皺,每個分子的面積不再發生真正的變化,盡管障礙物仍在移動。此外,由于褶皺,靠近探針的彎月面可能變得不規則,導致P測量中出現偽影。對于所有其他分子,可以觀察到膨脹軟化,表明軟分子被壓縮或部分分子從表面排出。因此,各層在壓縮后逐漸變弱。因此,在歧化過程中的某一點之后,隨著系統阻力的降低,氣泡收縮率將顯著增加。這與在工藝早期觀察到的情況完全相反,在該過程中,氣泡收縮引起的表面積小幅度減少會導致表面膨脹模量增加,從而加強該層并增加其抗歧化能力。此外,在大多數理論考慮中,未考慮此類行為,表面模量根據小變形實驗獲得的斜率或使用給定平衡吸附值的吉布斯彈性值作為常數。
從b-乳球蛋白的橢偏儀數據來看,后者確實如此(見圖8)。 從膨脹和剪切作用下的頻率行為可以看出,分子柔軟度和吸附能都很重要。
我們現在想將表面流變參數轉化為對泡沫中歧化穩定性的理解,并給出2Emax/gmax作為這一點的度量。 我們進一步注意到,根據Kloek等人的計算機模擬,在預測氣泡收縮時,2Emax/gmax應大于5而不是1.2,我們假設表面變形為純彈性。 然而,對于幾乎所有的材料,變形都是彈性和粘性作用的復雜量。
從頻率相關的表面膨脹和剪切流變學可以得出結論,HFBIIlayer的變形比Quillaja皂甙、b-乳球蛋白或b-酪蛋白層的變形更具彈性。 關于大變形、氣泡收縮和歧化,這表明HFBII的非平衡模量比其他材料的非平衡模量更受彈性貢獻的支配。
當我們現在再次考慮表1中的2Emax/gmax比率時,很明顯HFBII超過了5的值,這表明這將穩定泡沫,防止收縮。 根據Kloek等人的說法,對于b-乳球蛋白和皂甙,我們報告的值介于1和2之間,表明氣泡收縮延遲,但沒有完全停止。2食品蛋白質通常屬于這種狀態,對歧化過程的抵抗力相對較小。10最后, 根據這個比率,b-酪蛋白根本無法抵消氣泡收縮,因為2Emax/gmax<1,這是主要的吉布斯標準。
現在,我們將把表面流變學數據與泡沫的粗化時間以及早期報告9、25、26進行比較,其中探討了單個泡沫和泡沫的壽命。 我們發現HFBII穩定的泡沫或氣泡比其他所有泡沫或氣泡的壽命更長。 考慮到所研究表面層的全壓縮等溫線、2Emax/gmax比、松弛行為和表面剪切數據,我們發現表面膨脹實驗對氣泡抗收縮穩定性具有良好的預測能力。 更準確地說,可以看出,與其他系統相比,HFBII在所有膨脹量方面都非常突出,因此提供了一個獨特的機會,可以在更廣泛的泡沫壽命范圍內進行比較。 我們發現b-酪蛋白的最小粗化時間與它不符合吉布斯標準這一事實密切相關。 最后,b-lg和Quillaja皂甙的粗化時間相似,但它們比b-酪蛋白的粗化時間大。 同樣的趨勢也適用于表面膨脹參數,它們確實超過了吉布斯準則,但并不遠。 因此,這些物質可以減慢,但不能阻止歧化。
與表面膨脹流變學一樣,表面剪切模量似乎也給出了與tcoarse相同的趨勢。 然而,從理論上看,膨脹而非剪切是與氣泡收縮相關的主要變形。 因此,可以預期歧化率與膨脹模量之間存在直接聯系,而與剪切力之間存在間接聯系,因為通常所有表面參數都是表面吸附的相互依賴和單調函數。
最后,我們假設,如果一個吸附的蛋白質層既滿足2Emax/gmax>5的標準,又表現出有限的粘性損失,那么它可以在數月的時間尺度上有效地阻止歧化。
探索泡沫粗化與表面流變學之間的關聯性疏水性蛋白——摘要、介紹
探索泡沫粗化與表面流變學之間的關聯性疏水性蛋白——材料和方法
探索泡沫粗化與表面流變學之間的關聯性疏水性蛋白——結果和討論
探索泡沫粗化與表面流變學之間的關聯性疏水性蛋白——結論、致謝!