




合作客戶/
拜耳公司 |
同濟大學 |
聯合大學 |
美國保潔 |
美國強生 |
瑞士羅氏 |






相關新聞Info
應用不同組裝的磷脂酰膽堿對牛精漿蛋白的隔離:一種新的技術方法——結果和討論
來源:上海謂載 瀏覽 725 次 發布時間:2021-12-20
3.結果
3.1.精漿的界面性質
精漿SDS-PAGE電泳顯示存在一系列不同比例的14至120 kDa的多肽(圖1),與先前的研究結果一致[37]。在車道4上估計了14至30 kDa之間的五個主要波段的比例。
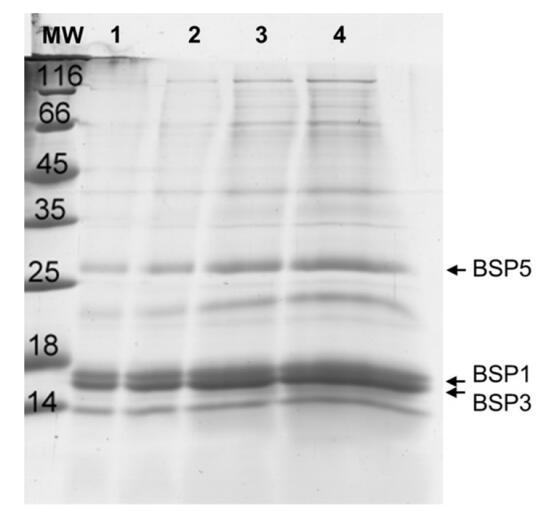
圖1。從牛射精中分離并以不同濃度沉積的精漿SDS-PAGE凝膠:lane 1:1μg;泳道2:2μg;泳道3:5μg和泳道4:10μg。
在大約16–17 kDa(74%強度)處發現了兩條非常緊密和強烈的條帶。它們被歸因于BSP1和BSP3[12]。在大約28 kDa(9%)處檢測到第二條帶,對應于BSP5[12]。BSP1、BSP3和BSP5代表精漿中所含最小多肽的83%(在整個提取物中約占65%)[17]。另一條帶在22kDa(4%)處檢測到,另一條帶在14kDa(13%)處檢測到,對應于核糖核酸酶[22]。
通過跟蹤精漿水溶液的表面張力(圖2)與濃度(吉布斯膜),突出了這些組分的表面活性。新樣品的表面張力相對較低(47 mN/m),并隨著樣品的稀釋而增加。在1/100000的大稀釋后,表面張力值等于純緩沖-空氣界面(72 mN/m)的值。因此,精漿顯示出顯著的表面活性:首先,新鮮樣品(47 mN/m)的表面張力與已知表面活性蛋白(如牛血清白蛋白[38]或溶菌酶[39])的值一樣低,其次,表面活性仍然存在于較大的濃度范圍內(需要稀釋100000以去除表面活性)。此外,通過朗繆爾薄膜分析了空氣-緩沖界面處表面薄膜的穩定性。薄膜的壓縮等溫線(圖3)顯示壓力在兩個步驟中從0顯著增加到35 mN/m。在18.9 mN/m時,出現斜率斷裂,這與空氣-緩沖界面分子構象的變化有關。在較高壓力下壓縮膜的能力(大約與生物膜相關的壓力,25 mN/m)表明,由精漿成分形成的膜具有較高的穩定性。
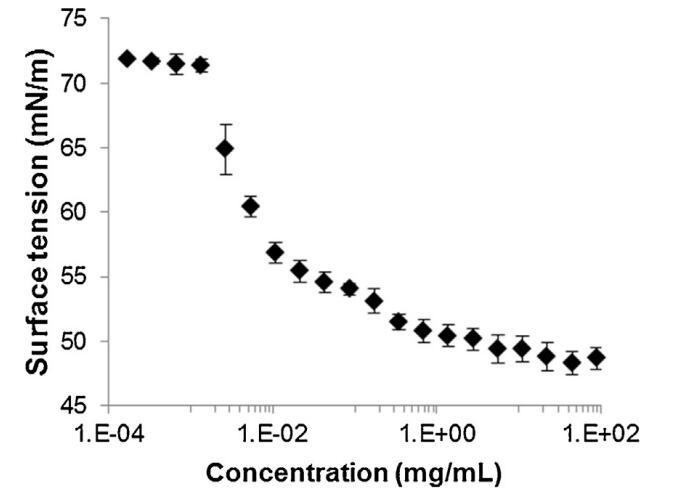
圖2。牛精漿溶液在空氣-水界面處的表面張力(±標準偏差),與提取物中所含蛋白質的濃度有關,并以對數標度表示(T=23.2℃±0.3℃,n=2,每個樣品重復4次)。
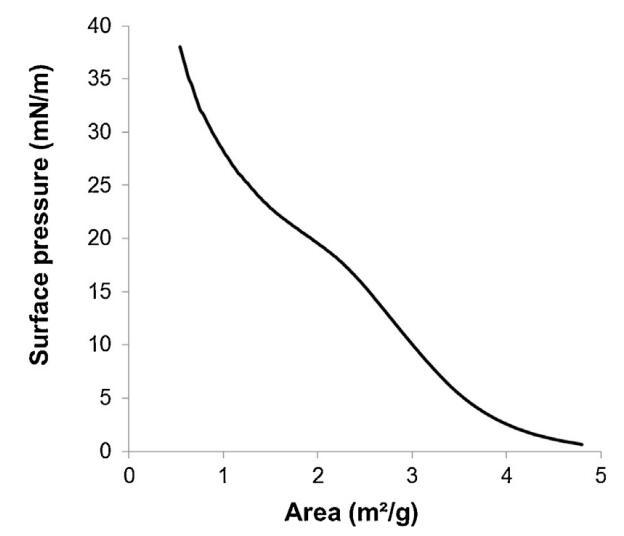
圖3。通過傳播精漿在空氣/緩沖液界面形成的單層的表面壓力面積(π–A)等溫線(n=3,標準偏差在0-18.9 mN/m范圍內不超過1.5 mN/m,在18.9-38 mN/m范圍內不超過1.2 mN/m)。
3.2.低密度脂蛋白和脂質體對精子膜仿生外小葉的作用
將低密度脂蛋白和脂質體引入精子膜仿生外小葉亞期的影響如圖4所示。對于控制測量(未注入任何保護劑,設計為圖4中的M),表面積隨時間略微減小。當低密度脂蛋白被注射到重建的外小葉下方的亞相(圖4中的曲線M/LDL)時,它導致表面積快速而大的增加。相反,與參考單層(曲線M)相比,脂質體的注射(圖4中的曲線M/C4)幾乎沒有改變單層的膜壓力。脂質體與單層脂質體的結合動力學比低密度脂蛋白慢得多。因此,在與精液制備相對應的時間尺度上,脂質體在空氣-水界面上的吸附較弱。
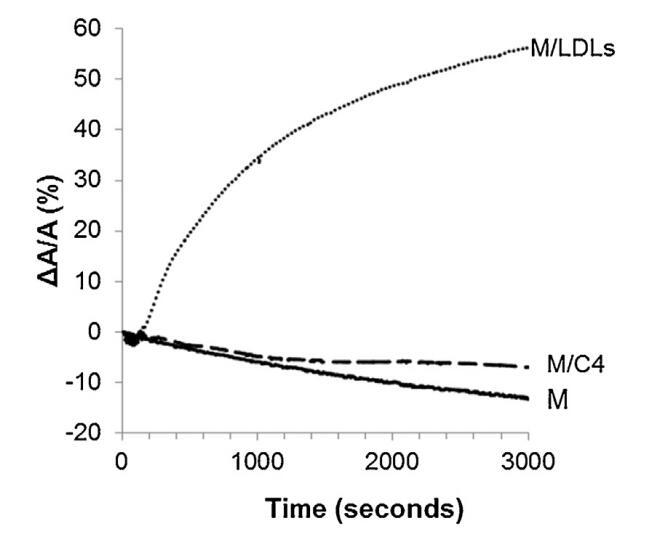
圖4。單層(M)所覆蓋的界面的相對表面積隨時間的變化,該單層(M)以25 mN/M的速度壓縮,并在不含任何額外分子(-)的緩沖液上形成,(…)LDL(4.6 mg PC)和(-)脂質體C4(含4.8 mg PC)。空氣-緩沖界面處的溫度為34℃。
對于低密度脂蛋白,根據低密度脂蛋白的組成,注射磷脂酰膽堿的量估計為4.6毫克[33]。就脂質體而言,該分散體含有6.55±0.02 mg脂質,即4.8 mg磷脂酰膽堿。在重建的精子膜外小葉下注射低密度脂蛋白后,表面積的大幅增加可能與低密度脂蛋白在空氣-水界面自發吸附和擴散的能力有關[33]。低密度脂蛋白在界面處的容易破壞可能釋放出磷脂和蛋白質,這些磷脂和蛋白質是表面活性物質,而甘油三酯是不可溶的[33]。對于脂質體,形成雙層磷脂和形成單層磷脂之間的平衡取決于鏈不飽和度、鏈長和極性頭基[40]。考慮到雞蛋卵磷脂(PC 74%)的主要成分及其脂肪酸組成[41],飽和PC和不飽和PC的比例分別約為48%和52%。在本研究的溫度下,飽和PC未熔化,可能不會在界面擴散,而不飽和PC遠遠超過其熔化溫度,并可能擴散。在剩余的部分PE(11%)中,形成磷脂的雙層和單層之間的平衡與磷脂的結構密切相關,因此不能對它們給出任何結論。因此,沒有基本的解釋。雞蛋脂質體缺乏吸附可以解釋為其擴散之前已在自由界面上進行過檢查:界面上已經存在的單層可能會限制磷脂基脂質體的擴散,而磷脂基脂質體在其他地方被認為是高穩定性的散裝物,尤其是由磷脂酰膽堿組成時。
3.3.低密度脂蛋白和脂質體分離精漿成分
將牛精漿(SP)注射到單層下方時,監測分子面積隨時間的變化(圖5中的曲線M/SP)。由于脂質覆蓋了界面,表面積并沒有立即增加,而是在150s后才增加。它在150秒內達到最大值(注射后300秒),然后穩定300秒并隨時間緩慢下降(圖5)。表面積的快速增加表明SP組分(很可能是蛋白質)具有良好的表面活性,如圖所示。2和3。600秒后,比表面積的減小速度與純單分子膜相同。因此,BSP蛋白一旦被吸附,就會留在界面上。
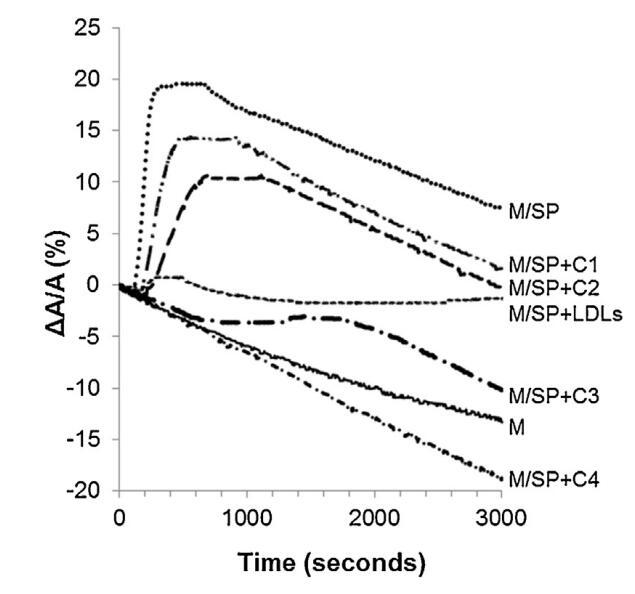
圖5。注入(…)后,由單層(M)覆蓋并在不含(-)額外分子的緩沖液上形成的界面的相對表面積隨時間的變化SP(26.6±1.5 mg蛋白質)、SP(26.6±1.5 mg蛋白質)和不同濃度(··-)C1、(·-)C2的脂質體的混合物,(·-)C3和(·-)C4(分別為1.17±0.02 mg、2.18±0.02 mg、4.37±0.02 mg和6.55±0.02 mg脂質)和(---)LDL(4.6 mg PC)和SP(26.6±1.5 mg蛋白質)的混合物。空氣-緩沖界面處的溫度為34℃。表面壓力保持在25 mN/m。
當LDL與牛精漿以重量-蛋白質比10:1(BSP:LDL)混合并注射到單層下方時,隨時間記錄的單層分子面積幾乎沒有變化(曲線M/SP+LDL,圖5)。400秒左右出現一個肩部,顯示BSP蛋白的弱吸附(該肩部代表M/SP的5%,上圖曲線)。然后,分子面積的變化在1000秒后減小并趨于穩定。其下降幅度不如M和M/SP曲線的變化幅度大,表明LDL組分吸附,與它們在界面上的高擴散能力一致(曲線M/LDL,圖4)。因此,由于低密度脂蛋白(LDL),SP中BSP蛋白在界面上的吸附大大減少,BSP/LDL的比率非常接近最佳值,以完全防止BSP蛋白影響單層。根據低密度脂蛋白的組成,注射磷脂酰膽堿的量估計為4.61毫克[33]。因此,向4.61 mg低密度脂蛋白磷脂酰膽堿注射26.6 mg精漿蛋白質,即0.17 mg磷脂酰膽堿注射1 mg精漿蛋白質。
脂質體(不同濃度)對BSP蛋白的作用也如圖5所示。脂質體在進入亞階段前與精漿接觸。隨著脂質體中濃度的增加(曲線M/SP+C1至C4),BSP蛋白吸附引起的表面積增加逐漸延遲并減少。當含量達到4.37 mg(C3)時,界面上仍有少量BSP蛋白摻入,1600 s時出現肩部。在最高濃度(C4)下,未觀察到BSP蛋白摻入,分子面積的變化甚至比對照曲線更負,表明膜組分在亞相中輕微溶解。因此,脂質體作用于BSP蛋白的方式與LDL相同。最有趣的比例似乎介于C3和C4之間。根據確定面積變化與脂質體注射磷脂質量之間相互作用的曲線計算得出(圖6)。注射26.6 mg BSP的最佳質量為4.6 mg,即PC:BSP比為0.16(w/w)。
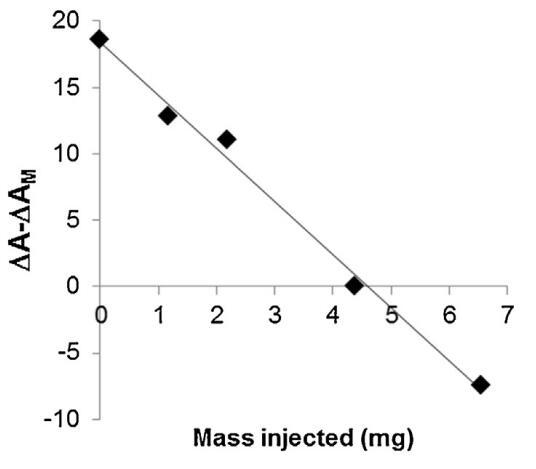
圖6。相對面積隨注射到亞相的脂質體質量的變化。線性回歸的R2為0.992。
3.4.脂質體對精漿作用的顯微評價
用透射電鏡(TEM)觀察脂質體或LDL與含有SP的BSP蛋白的混合物,并與對照組進行比較。無論放大倍數如何,與精漿相對應的圖像均未顯示任何結構(圖S1)。相反,與LDL和脂質體相關的結構很容易通過(i)LDL的致密且相當圓的形狀(圖7A)和(ii)脂質體的圓形且有時為多層結構(圖7B)來識別。在存在精漿的情況下,對整套圖像的分析仍然顯示出圓形物體。它們的大小平均與未使用螯合劑分析的大小相似(圖S2)。然而,在SP存在的幾個地方,觀察到一些大的圓形物體,其特征是(i)被厚層(圖7C含LDL)包圍的天然狀結構,或(ii)環形內的致密核心(圖7D含脂質體和圖S3)。與圖7B相比,與精漿混合的脂質體系統地呈現出比天然脂質體更致密的核心。這些新結構可能是由于BSP蛋白滲透到界面層而導致膜破裂的結果,從而導致TEM對比度的改變。

圖7。(A)低密度脂蛋白,(B)脂質體,(C)低密度脂蛋白和精漿(精漿中的比例為0.17 mg PC/mg BSP蛋白質)和(D)脂質體和精漿(精漿中的比例為0.16 mg PC脂質體/mg BSP蛋白質)的冷凍電鏡圖像。
4.討論
4.1.BSP蛋白的表面性質及其可能的生物學作用
張力測定結果顯示精漿中含有表面活性成分。這些很可能是蛋白質,因為它們構成精漿的主要部分。它們由幾個親水性和疏水性域組成[5],賦予它們兩親性。考慮到生物學背景,我們可以假設BSP蛋白能夠到達磷脂覆蓋的界面,這是由于它們與PC的相互作用和自身的表面活性。這可以解釋為什么這個過程很快[42]。將肽或蛋白質插入磷脂單分子膜通常在比凝聚域更可壓縮的流體域中進行[43]。在34°C時,液域由血漿鹵素和不飽和磷脂酰膽堿組成[35]。因此,根據前面提到的涉及疏水腔的機制,BSP蛋白可以很容易地穿透這些流體結構域,在這些流體結構域中它們可以立即與含有磷酸膽堿的分子相互作用[11,12,34,42,44–46]。吸附的BSP量可能受到流體域可壓縮性的限制。綜上所述,BSP蛋白質一旦被吸附,由于其表面性質(我們的工作)、構象重排[47]以及它們與膜組件的強烈相互作用[48],仍然固定在界面上。
4.2.脂質體和低密度脂蛋白在分離BSP蛋白中的作用
我們的結果表明,只要使用合適的脂質體/BSP比例,脂質體就可以像LDL一樣有效地阻止BSP蛋白到達界面。對于低密度脂蛋白,我們的模型顯示,低溫保存中常用的BSP/LDL比率(10:1 w/w)非常接近最佳值,以完全防止BSP蛋白影響單層。低密度脂蛋白組織中PC/BSP與PC的比率為0.17 mg磷脂酰膽堿,1 mg精漿蛋白質。同樣,脂質體的最佳比例PC/BSP為0.16 mg磷脂酰膽堿和1 mg精漿蛋白質。知道卵子中PC的比例可能取決于喂食情況,不同批次的LDL中PC的比例可能不同,因此這些比例被認為是相等的。該比率與LDL或脂質體的隔離結構無關的事實表明,LDL中的載脂蛋白不與BSP結合,盡管這種結合是可能的[26,49]。更引人注目的是,這一結果表明PC作為LDL中的單層或脂質體中的雙層組裝沒有影響。在低密度脂蛋白的情況下,磷脂占據外層單層,并且所有這些磷脂都可被蛋白質接近。相反,只有形成單層脂質體的雙層外小葉的磷脂真正與BSP蛋白直接接觸。由于脂質體和低密度脂蛋白的絡合比率相同,并且PCP分子(74%)也構成雙層的內小葉,因此這些分子也參與BSP蛋白的絡合。與BSP蛋白易于插入疏水膜相一致,這些BSP蛋白可向內小葉遷移,并誘導脂質體雙層破裂/融合,如TEM所觀察到的。然而,樣品中LDL和脂質體的結構變化并不普遍,BSP蛋白可以認為主要錨定在LDL和脂質體的表面。當脂質體與BSP的比例高于本文所研究的比例時,BSP1(BSP家族中的主要蛋白質)誘導脂質小泡的伸長和珠項鏈狀結構的形成,這些結構演變為小泡或線狀結構[50]。